
February 6, 2018
Adipokines and Adipose Tissue-Liver Crosstalk in Energy Metabolism
Research over the past several decades examining adipokines and adipose tissue-liver crosstalk in energy metabolism has revealed that adipose tissue is a very important metabolically active endocrine organ1. White, brown, and beige adipose tissue generate and secrete adipokines, such as adiponectin and leptin, which facilitate adipose tissue-liver crosstalk and insulin sensitivity1.
https://www.youtube.com/watch?v=_1il8nV5V1Q&feature=youtu.be
White adipose tissue (WAT) contains white adipocytes and is the most common type found in adult humans3,5. White adipocytes contain a single large vacuole that stores lipids produced via lipogenesis and lipids from food consumed6. WAT can expand either through size (hypertrophy) or number (hyperplasia) to accommodate excessive lipid levels7.
There are two types of WAT found in different areas of the body. Subcutaneous white adipose tissue (SWAT) is found under the skin throughout the body and has several functions including acting as a layer of insulation1. Visceral white adipose tissue (VWAT) is located around the internal organs and is the main type of adipose tissue used for energy storage. VWAT also offers organs mechanical protection3. The abdominal cavity usually contains a large VWAT depot1.
Research indicates that the different SWAT and VWAT deposits have varying gene expression profiles and, therefore, contribute to energy balance in different ways. VWAT has been shown to secrete more inflammatory cytokines as compared to SWAT. Studies demonstrate that SWAT expresses higher levels of transcription factors that promote preadipocyte differentiation6.
Brown Adipose Tissue
Brown adipose tissue (BAT) contains brown adipocytes. Unlike white adipocytes, brown adipocytes contain many small vacuoles that hold lipids3,5. Additionally, brown adipocytes are abundant in mitochondria which contribute to the brown appearance of these fat cells3. The main function of BAT is to oxidize lipids for heat production through a process known as non-shivering thermogenesis3,5,7.
Research has shown that BAT is more bountiful during development and early stages of life. Then with age, BAT stores are lost and WAT stores increase3,5. Experiments using positron emission tomography (PET) demonstrated that BAT is still present in human adults in small amounts, mainly beneath the neck and in between the shoulders3.
 White, brown, and beige adipose tissue perform highly regulated metabolic functions3. Adipocytes in adipose tissue secrete many cell signaling molecules including adipokines which respond to different stimuli and communicate with other organs1,3,8. Adiponectin and leptin are two central adipokines that interact with the liver to facilitate energy balance4,8,19. Therefore, secreted adipokines and adipose tissue-liver crosstalk in energy metabolism regulation are essential for maintaining overall homeostasis4,8 .
White, brown, and beige adipose tissue perform highly regulated metabolic functions3. Adipocytes in adipose tissue secrete many cell signaling molecules including adipokines which respond to different stimuli and communicate with other organs1,3,8. Adiponectin and leptin are two central adipokines that interact with the liver to facilitate energy balance4,8,19. Therefore, secreted adipokines and adipose tissue-liver crosstalk in energy metabolism regulation are essential for maintaining overall homeostasis4,8 .
What is Adipose Tissue?
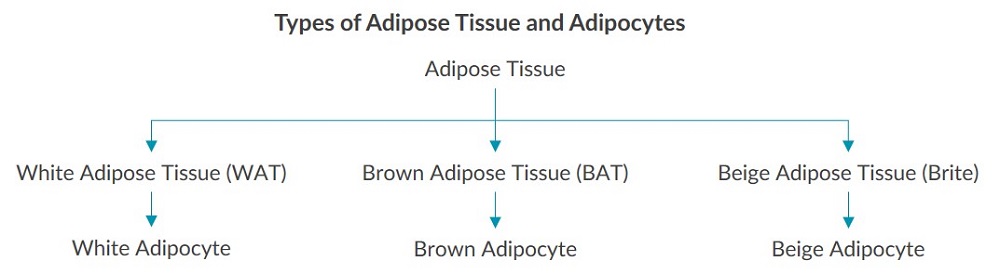
Adipose tissue (AT), also referred to as fat tissue, is found throughout the body and is vital for maintaining energy homeostasis1. Several cell types are found in adipose tissue; however, the main type is the adipocyte, commonly known as a fat cell. Adipocytes take up 90% of the volume in adipose tissue2. White, brown and beige adipose tissue each contain a corresponding type of adipocyte as depicted below3:
Functions of Adipose Tissue
Each type of adipose tissue and adipocyte contributes differently to energy metabolism and overall homeostasis3. One way adipose tissue supports energy metabolism is through the generation and secretion of pro- and anti-inflammatory cytokines and hormones. Cytokines and hormones exclusively produced by adipose tissue are referred to as adipokines. Each type of AT releases a different combination of these cell signaling molecules which enable the highly regulated crosstalk between adipose tissue and other organs such as the liver and brain to maintain energy balance4. The function of adipose tissue not only depends on the type of adipocyte, but the location of the fat deposit as well. Additionally, the environment, genetics, age, gender, nutritional status, and weight can impact the structure and function of each type of adipose tissue2,3. Due to these differences, some researchers refer to each site of adipose tissue as a mini organ, contributing differently to energy metabolism2. White Adipose Tissue
White Adipose Tissue
White adipose tissue (WAT) contains white adipocytes and is the most common type found in adult humans3,5. White adipocytes contain a single large vacuole that stores lipids produced via lipogenesis and lipids from food consumed6. WAT can expand either through size (hypertrophy) or number (hyperplasia) to accommodate excessive lipid levels7.
There are two types of WAT found in different areas of the body. Subcutaneous white adipose tissue (SWAT) is found under the skin throughout the body and has several functions including acting as a layer of insulation1. Visceral white adipose tissue (VWAT) is located around the internal organs and is the main type of adipose tissue used for energy storage. VWAT also offers organs mechanical protection3. The abdominal cavity usually contains a large VWAT depot1.
Research indicates that the different SWAT and VWAT deposits have varying gene expression profiles and, therefore, contribute to energy balance in different ways. VWAT has been shown to secrete more inflammatory cytokines as compared to SWAT. Studies demonstrate that SWAT expresses higher levels of transcription factors that promote preadipocyte differentiation6.
Brown Adipose Tissue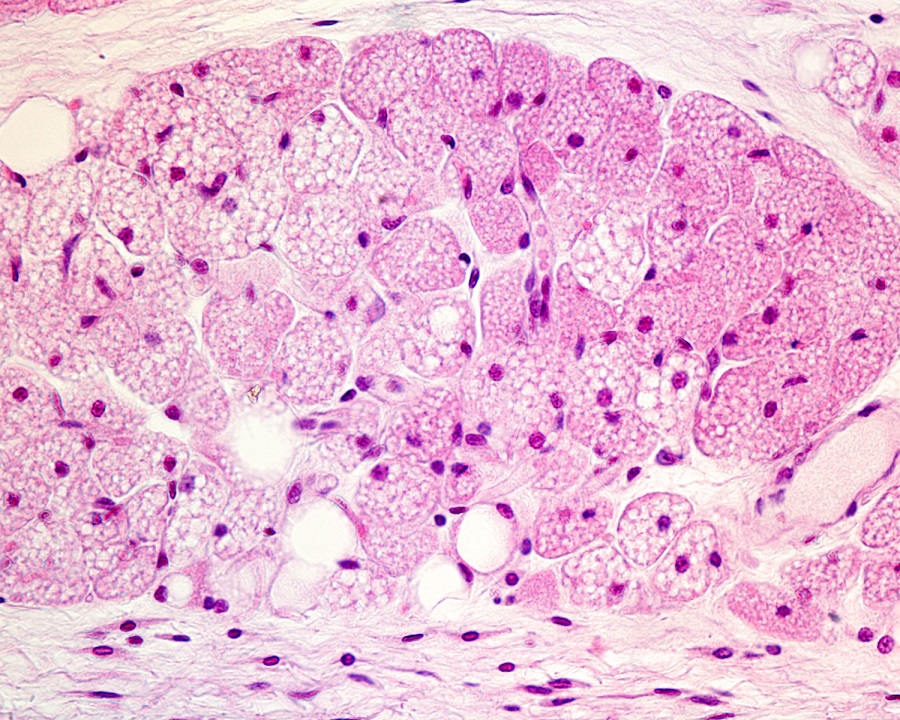
Brown adipose tissue (BAT) contains brown adipocytes. Unlike white adipocytes, brown adipocytes contain many small vacuoles that hold lipids3,5. Additionally, brown adipocytes are abundant in mitochondria which contribute to the brown appearance of these fat cells3. The main function of BAT is to oxidize lipids for heat production through a process known as non-shivering thermogenesis3,5,7.
Research has shown that BAT is more bountiful during development and early stages of life. Then with age, BAT stores are lost and WAT stores increase3,5. Experiments using positron emission tomography (PET) demonstrated that BAT is still present in human adults in small amounts, mainly beneath the neck and in between the shoulders3.
Beige Adipose Tissue
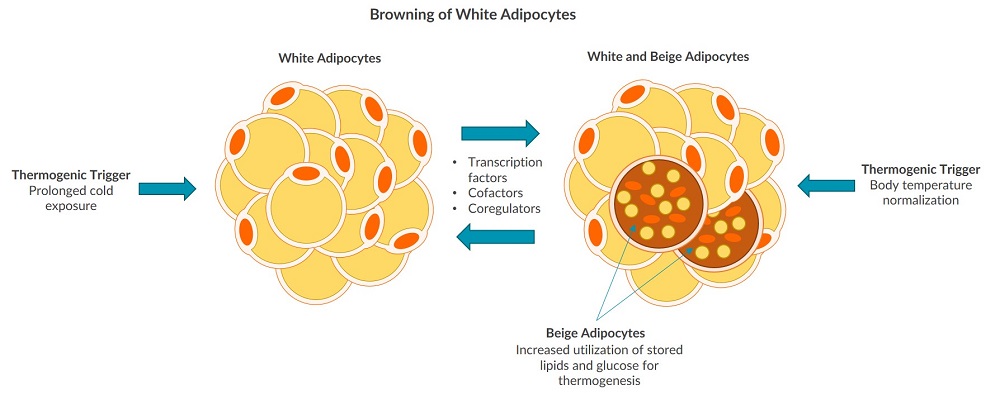
The third type of adipose tissue found in humans is beige adipose tissue. Beige adipose tissue is found scattered throughout SWAT deposits and does not have its own dedicated fat pads1,5. Compared to WAT, beige adipose tissue has more mitochondria and performs more fatty acid oxidation, but stores less lipids9. The adipocytes found in beige adipose tissue are commonly referred to as beige or brite adipocytes. Brite is derived from the phrase brown in white due to the ability of WAT to undergo a process referred to as browning5. White adipose tissue browning can be initiated by a thermogenic stimulus, such as prolonged exposure to cold5. It has been shown that beige adipocytes have the ability to revert back to white adipocytes once the body reaches normal temperature again, then back to beige adipocytes upon more prolonged cold exposure1. Physical exercise, interleukins, noncoding micro RNAs, proteins, and epigenetic changes may also affect WAT browning9. Researchers are working to fully understand if beige adipocytes arise from the same precursor cells as white adipocytes, or from their own distinctive precursors5.
Functions of Adipokines in Energy Metabolism
White, brown, and beige adipose tissue synthesize and release adipokines to influence energy metabolism8. Adiponectin (ADP) and leptin are two key adipokines secreted by adipose tissue. Adiponectin and leptin each perform functions to support homeostasis and are essential for proper adipose tissue-liver crosstalk. Due to their roles in energy homeostasis, both adiponectin and leptin have become cornerstone biomarkers in metabolic research4.Adiponectin
Adiponectin is a 244 amino acid hormone exclusively synthesized and secreted by adipose tissue10. This adipokine plays many different physiological roles such as promoting insulin sensitivity and regulating lipid metabolism10. Research indicates that insulin promotes the release of ADP into circulation, while the proinflammatory cytokine TNF-alpha inhibits the adipokine11. SWAT has been shown to secrete higher concentrations of adiponectin than VWAT8.Leptin
Leptin was discovered as the first adipokine in 1994 and its finding demonstrated that adipose tissue is an active endocrine organ12,13. WAT secretes leptin into circulation as a 16kDa protein that controls food intake and regulates energy homeostasis by signaling fullness to the brain14,15. Due to its role in facilitating satiety, leptin is also known as a hunger hormone16. Overall levels of leptin are related to the amount of adipose tissue, current triglyceride energy stores, and changes to energy intake13,17. Research shows that SWAT secretes higher levels of leptin than VWAT, and that leptin levels are higher in women8.Adipokines and Adipose Tissue-Liver Crosstalk in Energy Metabolism
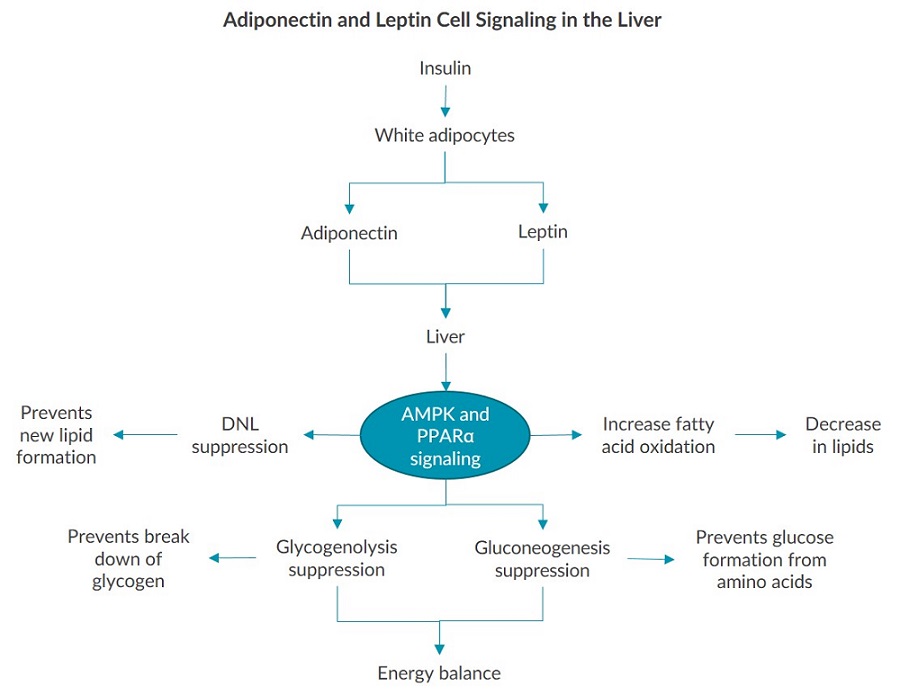
Research over the past 20 years demonstrates how adiponectin and leptin are key components of the adipose tissue-liver crosstalk that regulates energy metabolism4. The liver expresses the leptin receptor and all three adiponectin receptors. This expression allows the adipokines to act as messengers from adipose tissue to the liver, influencing glucose uptake, lipid oxidization and de novo lipogenesis (DNL)4,10,18.Adipokine Cell Signaling in the Liver
The cell signaling pathways in the liver triggered by adiponectin and leptin are mandatory for regulating energy metabolism. Both adipokines activate AMP protein kinase (AMPK) which inhibits enzymes and transcription factors needed for liver de novo lipogenesis4. Additionally, adiponectin can activate peroxisome proliferator-activated receptor (PPARα) which also suppresses DNL4. AMPK and PPARα signaling in the liver is triggered by leptin and adiponectin binding to their receptors, thus influencing glucose metabolism by suppressing glycogenolysis and gluconeogenesis. In turn, the levels of both glucose and fatty acids can remain in homeostatic range as a result of leptin and adiponectin interacting with the liver4,19.
White, brown, and beige adipose tissue perform highly regulated metabolic functions3. Adipocytes in adipose tissue secrete many cell signaling molecules including adipokines which respond to different stimuli and communicate with other organs1,3,8. Adiponectin and leptin are two central adipokines that interact with the liver to facilitate energy balance4,8,19. Therefore, secreted adipokines and adipose tissue-liver crosstalk in energy metabolism regulation are essential for maintaining overall homeostasis4,8 .
References
- Choe, et al (2016). Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Metabolism and Metabolic Disorders. Front. Endocrinol. 7:30. PMCID: PMC4829583.
- Badimon and Cubedo (2017). Adipose Tissue Depots and inflammation: effects on plasticity and resident mesenchymal stem cell function. Cardiovasc Res. 2017 Jul 1;113(9):1064-1073. PMID: 28498891.
- Esteve Rafol (2014). Adipose Tissue: Cell Heterogeneity and Functional Diversity. Endocrinol Nutr. 2014 Feb;61(2):100-12. PMID: 23834768.
- Stern, et al (2016). Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016 May 10;23(5):770-84. PMID: 27166942.
- Giralt and Villarroya (2013). White, Brown, Beige/Brite: Different Adipose Cells for Different Functions? Endocrinology. 2013 Sep;154(9):2992-3000. PMID: 23782940.
- Cleal, et al (2017). Fifty Shades of White: Understanding Heterogeneity in White Adipose Stem Cells. 2017; 6(3): 205–216. PMCID: PMC5638386.
- Pellerginelli, et al (2016). Adipose Tissue Plasticity: How Fat Depots Respond Differently to Pathophysiology. Diabetologia. 2016 Jun;59(6):1075-88. PMID: 27039901.
- Coelho, et al (2013). Biochemistry of adipose tissue: an endocrine organ. Arch Med Sci. 2013 Apr 20; 9(2): 191–200. PMCID: PMC3648822.
- Lizcano and Vargas (2016). Biology of Beige Adipocyte and Possible Therapy for Type 2 Diabetes and Obesity. Int J Endocrinol. 2016;2016:9542061. PMID: 27528872.
- Wang and Scherer (2016). Adiponectin, the past two decades. J Mol Cell Biol. 2016 Apr;8(2):93-100. doi: 10.1093/jmcb/mjw011. PMID: 26993047
- Wang, et al (2007). Secretion of the Adipocyte-Specific Secretory Protein Adiponectin Critically Depends on Thiol-Mediated Protein Retention. Mol. Cell. Biol. May 2007 vol. 27 no. 10 3716-3731. doi: 10.1128/MCB.00931-06.
- Zhang, et al (1994). Positional cloning of the mouse obese gene and its human homologue. Nature 372 425–432. doi:10.1038/372425a0.
- Polyzos, et al (2016). Circulating Leptin in non-alcoholic fatty liver disease: a systemic review and meta-analysis. Diabetologia. 2016 Jan;59(1):30-43. PMID: 26407715.
- Myers, et al (2008). Mechanisms of leptin action and leptin resistance. Rev. Physiol., 70, 537-56. PMID: 17937601.
- Park and Ahima (2015). Physiology of leptin: Energy homeostasis, neuroendocrine function and metabolism. Metabolism Clinical and Experimental, 64, 24-34.1. PMID: 25199978.
- Klok, et al (2007). The role of leptin and ghrelin in the regulation of food intake and body weight in humans: a review. Obes Rev. 2007 Jan;8(1):21-34. PMID: 17212793.
- Meyers, et al (2010). Obesity and Leptin Resistance: Distinguishing Cause from Effect. Trends Endocrinol Metab. 2010 Nov; 21(11): 643–651. PMCID: PMC296765.
- Ghantous, et al (2015). Differential Role of Leptin and Adiponectin in Cardiovascular System. International Journal of Endocrinology. Volume 2015 (2015),
- Article ID 534320,Combs and Marliss (2014). Adiponectin Signaling in the Liver. Rev Endocr Metab Disord. 2014 Jun;15(2):137-47. doi: 10.1007/s11154-013-9280-6. PMID: 24297186